همانند سازی DNA
همانندسازی DNA
زمانی که مولکول DNA برای اولین بار شناخته شد، یکی از خصوصیاتی که توجه همگان را به خود جلب کرد ویژگی مکمل بودن زنجیرههای آن بود. کشف ساختمان فوق ایدهای در مورد همانند سازی DNA در اختیار قرار میداد. ضمناً این کشف دانشمندان را بر آن داشت تا نظر اوری که مولکول DNA (نه پروتئینها) حملکننده اطلاعات ژنتیکی است را قبول کنند.
در بحثهای قبلی اشاره شد که در ساختمان مکمل سطوح مشابه نمیتوانند همدیگر را جذب کنند و لازمه جذب، وجود شکل و بار الکتریکی مخالف در دو ساختمان است (منظور از شکل مخالف این است که اگر در یک زنجیره برآمدگی وجود دارد، در زنجیره مقابل باید فرورفتگی وجود داشته باشد تا ساختمان مکمل تشکیل شود). بنابراین بدون در دست داشتن اطلاعات ساختمانی میتوان حدس زد که عمل کپیبرداری از یک ژن نمیتواند مستقیم و بدون واسطه باشد، بلکه در واقع مولکولی که مکمل ژن اصلی است به عنوان الگو قرار میگیرد و از روی آن مولکولی شبیه ژن اصلی ساخته میشود. قبل از اینکه اطلاعات کافی در مورد اسیدهای نوکلئیک و پروتئینها در دسترس ژنتیکدانان باشد، تصور بر این بود که مولکول DNA برای بیوسنتز پروتئینها، خود به عنوان الگو قرار میگیرد و پروتئین تولید شده نیز به عنوان الگو در سنتز DNA شرکت میکند.
ولی بعد از آنکه معلوم شد که دو رشته DNA مکمل یکدیگر میباشند، نقش پروتئین به عنوان الگو در همانندسازی DNA رد گردید. در عوض گفته شد که هر یک از رشتههای DNA مادر به عنوان الگو قرار گرفته و مولکولهای DNA دختر از روی آنها بوجود میآیند. اگر چه از همان ابتدا فرضیه فوق بسیار منطقی بنظر میرسید، با این حال شواهد لازم برای اثبات آن باید فراهم میگشت. خوشبختانه پنجسال پس از شناخت ساختمان مارپیچ مضاعف، مشاهده شد که در هنگام همانندسازی دو زنجیره مکمل به طور موقت از یکدیگر جدا میشوند و شواهد آنزیمی موثق دال بر این بود که مولکول DNA در هنگام همانندسازی به عنوان الگویی جهت ساخت زنجیرههی جدید عمل میکند.
یافتههای فوق مسئله همانند سازی ژن را تا حدودی روشن کرد ولی مطالعات وسیعتر در مورد جزئیات همانندسازی DNA قدمهای نخستین را میپیمود. همانندسازی DNA فرایند چندمرحلهای و پیچیدهای است. به طوری که همانندسازی سادهترین مولکولهای DNA نیز به آنزیمهای متعددی نیاز دارد، در حالی که در ابتدا تصور میشد که این عمل تنها به وسیله یک آنزیم پلیمریزه کننده صورت میگیرد. خوشبختانه کلیه آنزیمهایی که در همانندسازی مولکولهای خطی و حلقوی DNA شرکت میکنند شناخته شدهاند و تصویری که در این فصل ارائه میشود در آینده تغییر چندانی نخواهد کرد.
جدا شدن زنجیرههای DNA مستلزم باز شدن تابهای مارپیچ مضاعف است.
همانگونه که پیشتر گفته شد، DNA به صورت مارپیچ مضاعفی است که در آن دو رشته به دور یکدیگر تاب خوردهاند و برای انجام همانندسازی، تابهای فوق باید باز شوند. چگونگی باز شدن این تابها ابتدا برای برخی از دانشمندان بسیار پیچیده بنظر میرسید ولی اگر قسمتهایی که هنور همانندسازی نکردهاند بتوانند حول محور خود بچرخند باز شدن تاب دو زنجیره آنچنان مشکلی را بوجود نمیآورد(شکل).
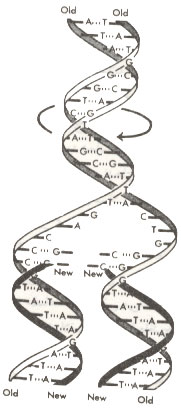
| باز شدن پیچهای DNA خطی در هنگام همانندسازی. زنجیرهها با چرخش به دور محور مارپیچ باز میشوند. |
ضمناً از آنجا که مولکولهای DNA بسیار باریک میباشند باز شدن تابهای فوق اصطکاکی بوجود نمیآورد و برای انجام آن انرژی ناچیزی مورد نیاز است.
شکسته شدن پیوندهای هیدروژنی که دو زنجیره را در کنار یکدیگر قرار میدهند، آنچنان مشکل نیست، چرا که با وجودی که این پیوندها به طور اختصاصی بوجود آمدهاند ولی نسبتاً ضعیف هستند و برای تشکیل و قطع آنها به آنزیمی احتیاج نیست. در واقع پیوندهای هیدروژنی سبب کنار هم قرار گرفتن الگو و رشته مکمل آن میشوند.
جفت شدن بازها امکان انجام دقیق همانندسازی را فراهم میسازد
در مطالب قبل اشاره شد که خواص گروههای جانبی اسیدهای آمینه مانع از بکارگیری آنها به عنوان الگوست ولی این موضوع در مورد بازهای پورینی و پیریمیدینی صدق نمیکند، چرا که آنها میتوانند پیوندهای هیدروژنی متعددی بوجود آورند و از آنجا که پیوندهای فوق کاملاً اختصاصی هستند، اسیدهای نوکلئیک میتوانند به عنوان الگو محسوب شوند. یادآوری میگردد که پیوندهای واندروالسی که بین گروههای جانبی اسیدهای آمینه میتوانند بوجود آیند بسیار ضعیف میباشند، ضمن اینکه ایجاد این پیوندها وابسته به وجود گروه شیمیایی خاصی نیست.
متوسط انرژی یک پیوند هیدروژنی حدود 3 کیلوکالی بر مول است که تقریباً هشت برابرانرژی حرارتی ناشی از حرکت مولکولها در دمای اتاق است. عدد فوق نشان میدهد که تا چه حد گروههای مختلف (مثل گروه آمین) میتوانند پیوند هیدروژنی ایجاد کنند.در شرایط سلولی نسبت پیوندهای ایجاد شده به حالت آزاد (حالت غیرپیوندی) 1 به 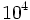 است. به عبارت دیگر اگر دو مولکول به وسیله چند پیوند هیدروژنی به یکدیگر متصل شوند تقریباً هیچگاه به صورت آزاد یافت نمیگردند. به دلیل اختصاصی بودن پیوندهای هیدروژنی در ساختمان مارپیچ مضاعف احتمال اتصال آدنین به سیتوزین هنگام همانندسازی
است. به عبارت دیگر اگر دو مولکول به وسیله چند پیوند هیدروژنی به یکدیگر متصل شوند تقریباً هیچگاه به صورت آزاد یافت نمیگردند. به دلیل اختصاصی بودن پیوندهای هیدروژنی در ساختمان مارپیچ مضاعف احتمال اتصال آدنین به سیتوزین هنگام همانندسازی  برابر کمتر از اتصال آن به تیمین است (
برابر کمتر از اتصال آن به تیمین است ( چون دو پیوند هیدروژنی بین آدنین و تیمین وجود دارد). همانند سازی جفتهای GC دقیقتر از AT است، چون C و G به وسیله سه پیوند هیدروژنی به یکدیگر متصل میشوند، با این حال به دلیل وجود توتومرهای غلط (ایمین یا انول) ممکن است جفتهای غلط انتخاب گردند (احتمال
چون دو پیوند هیدروژنی بین آدنین و تیمین وجود دارد). همانند سازی جفتهای GC دقیقتر از AT است، چون C و G به وسیله سه پیوند هیدروژنی به یکدیگر متصل میشوند، با این حال به دلیل وجود توتومرهای غلط (ایمین یا انول) ممکن است جفتهای غلط انتخاب گردند (احتمال  ) در نتیجه پس از همانند سازی تنها در اثر تصحیح میتوان صحت همانندسازی را تا حد قابل قبولی (
) در نتیجه پس از همانند سازی تنها در اثر تصحیح میتوان صحت همانندسازی را تا حد قابل قبولی (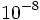 تا
تا 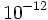 خطا) بالا برد.
خطا) بالا برد.
است. به عبارت دیگر اگر دو مولکول به وسیله چند پیوند هیدروژنی به یکدیگر متصل شوند تقریباً هیچگاه به صورت آزاد یافت نمیگردند. به دلیل اختصاصی بودن پیوندهای هیدروژنی در ساختمان مارپیچ مضاعف احتمال اتصال آدنین به سیتوزین هنگام همانندسازی برابر کمتر از اتصال آن به تیمین است (چون دو پیوند هیدروژنی بین آدنین و تیمین وجود دارد). همانند سازی جفتهای GC دقیقتر از AT است، چون C و G به وسیله سه پیوند هیدروژنی به یکدیگر متصل میشوند، با این حال به دلیل وجود توتومرهای غلط (ایمین یا انول) ممکن است جفتهای غلط انتخاب گردند (احتمال ) در نتیجه پس از همانند سازی تنها در اثر تصحیح میتوان صحت همانندسازی را تا حد قابل قبولی ( تا خطا) بالا برد.همچنین یکی از مسائلی که در مورد همانندسازی مطرح شد چگونگی تشکیل پیوندهای هیدروژنی بین بازهای آلی و مولکول آب بود. پیوندهای هیدروژنی بین آب و بازها (مثلاً تیمین) اهمیت چندانی ندارند زیرا کمپلکسهای فوق موقتی هستند. مولکولهای آب به زنجیرههای در حال رشد پلینوکلئوتیدی متصل نمیگردند و با اضافه شدن بازهای آلی مناسب کنار گذاشته میشوند. در زنجیره DNA هیچ گاه دو پورین یا دو پیریمیدین به وسیله پیوندهای هیدروژنی به یکدیگر متصل نمیشوند. تشکیل جفتهای پورین پورین و یا پیریمیدین پیریمیدین، ساختمان اسکلت اصلی مارپیچ مضاعف را عوض میکند و از نظر انرژی آرایش ناپایداری بوجود میآورد. با این حال محاسبه اختلاف انرژی به لحاظ حضور اتمهای متعدد بسیار دشوار است. یکی از علل دشواری محاسبات انرژی به دلیل مشکلات موجود در مشخص کردن مکان هر یک از اتمها با دقت یکدهم انگستروم است. همچنین مشخص نیست که تغییرات حاصله از تشکیل جفتهای پورین پورین و یا پیریمیدین پیریمیدین چه تاثیری در تشکیل آنزیمی پیوندهای کووالانسی بین نوکلئوتیدها میگذارد.
+ نوشته شده در دوشنبه بیست و پنجم آذر ۱۳۸۷ ساعت 20:53 توسط امین
|